
IPTG (isopropyl-β-D-thiogalactoside) เป็นสารอะนาล็อกของสารตั้งต้นของ β-galactosidase ซึ่งสามารถเหนี่ยวนำได้สูง ภายใต้การเหนี่ยวนำของ IPTG สารเหนี่ยวนำสามารถสร้างสารเชิงซ้อนกับโปรตีนยับยั้ง ทำให้โครงสร้างของโปรตีนยับยั้งเปลี่ยนแปลงไป ส่งผลให้ไม่สามารถรวมตัวกับยีนเป้าหมายได้ และยีนเป้าหมายจึงแสดงออกได้อย่างมีประสิทธิภาพ ดังนั้นควรพิจารณาความเข้มข้นของ IPTG ในการทดลองอย่างไร? ยิ่งมากยิ่งดีหรือไม่?
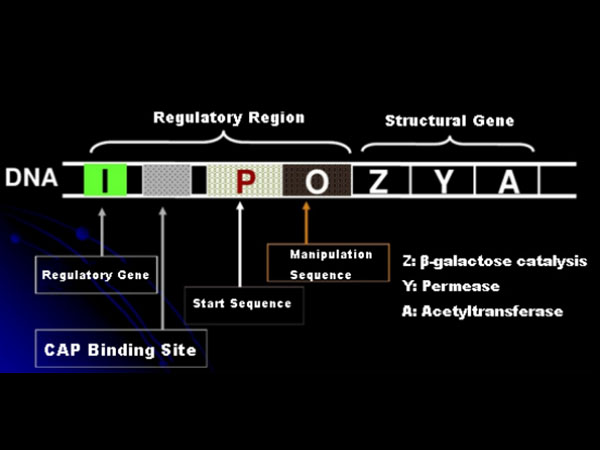
ก่อนอื่น เรามาทำความเข้าใจหลักการของการเหนี่ยวนำด้วย IPTG กันก่อน: โอเปรอนแลคโตส (องค์ประกอบ) ของ E. coli ประกอบด้วยยีนโครงสร้างสามยีน ได้แก่ Z, Y และ A ซึ่งเข้ารหัสเอนไซม์ β-galactosidase, permease และ acetyltransferase ตามลำดับ lacZ ทำหน้าที่ไฮโดรไลซ์แลคโตสให้เป็นกลูโคสและกาแลคโตส หรือเป็นอัลโล-แลคโตส; lacY ช่วยให้แลคโตสในสิ่งแวดล้อมผ่านเยื่อหุ้มเซลล์และเข้าสู่เซลล์; lacA ทำหน้าที่ถ่ายโอนหมู่แอซิทิลจากแอซิทิล-CoA ไปยัง β-galactoside ซึ่งเกี่ยวข้องกับการกำจัดผลกระทบที่เป็นพิษ นอกจากนี้ยังมีลำดับการทำงาน O, ลำดับเริ่มต้น P และยีนควบคุม I ยีน I เข้ารหัสโปรตีนตัวยับยั้งที่สามารถจับกับตำแหน่ง O ของลำดับการทำงาน ทำให้โอเปรอน (meta) ถูกยับยั้งและปิดการทำงาน นอกจากนี้ ยังมีตำแหน่งจับยึดของโปรตีนกระตุ้นยีนแคตาโบลิก (CAP binding site) อยู่เหนือลำดับเริ่มต้น P ลำดับ P, ลำดับ O และตำแหน่งจับยึด CAP รวมกันเป็นบริเวณควบคุมของโอเปรอนแลค ยีนที่เข้ารหัสเอนไซม์ทั้งสามชนิดถูกควบคุมโดยบริเวณควบคุมเดียวกันนี้ เพื่อให้เกิดการแสดงออกของผลิตภัณฑ์ยีนอย่างประสานกัน
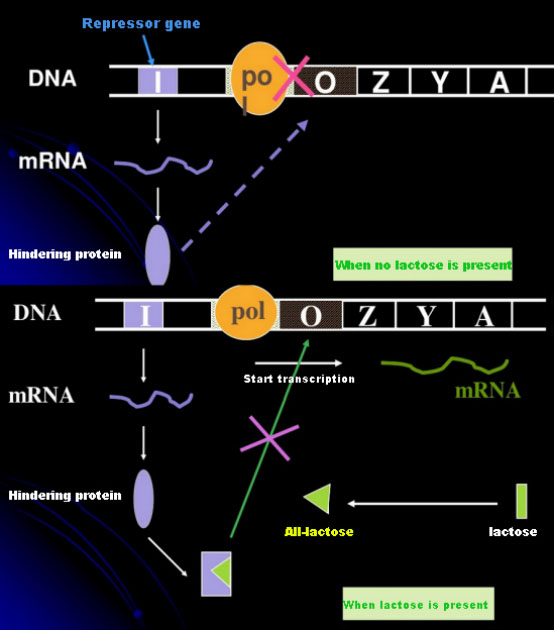
ในกรณีที่ไม่มีแลคโตส โอเปรอนแลค (เมตา) จะอยู่ในสภาวะถูกยับยั้ง ในขณะนี้ โปรตีนยับยั้งแลคที่แสดงออกโดยลำดับ I ภายใต้การควบคุมของลำดับโปรโมเตอร์ PI จะจับกับลำดับ O ซึ่งจะป้องกันไม่ให้ RNA โพลีเมอเรสจับกับลำดับ P และยับยั้งการเริ่มต้นการถอดรหัส เมื่อมีแลคโตส โอเปรอนแลค (เมตา) สามารถถูกกระตุ้นได้ ในระบบโอเปรอน (เมตา) นี้ ตัวกระตุ้นที่แท้จริงไม่ใช่แลคโตสเอง แลคโตสเข้าสู่เซลล์และถูกเร่งปฏิกิริยาโดย β-galactosidase เพื่อเปลี่ยนเป็นอัลโลแลคโตส อัลโลแลคโตสซึ่งเป็นโมเลกุลตัวกระตุ้น จะจับกับโปรตีนยับยั้งและเปลี่ยนโครงสร้างของโปรตีน ซึ่งนำไปสู่การแยกตัวของโปรตีนยับยั้งออกจากลำดับ O และการถอดรหัส ไอโซโพรพิลไทโอแกลแลคโตไซด์ (IPTG) มีผลเช่นเดียวกับอัลโลแลคโตส เป็นสารกระตุ้นที่มีประสิทธิภาพสูงมาก แบคทีเรียไม่สามารถย่อยสลายได้ และมีความเสถียรสูง จึงมีการใช้กันอย่างแพร่หลายในห้องปฏิบัติการ
จะกำหนดความเข้มข้นที่เหมาะสมที่สุดของ IPTG ได้อย่างไร? ลองใช้ E. coli เป็นตัวอย่าง
สายพันธุ์ E. coli BL21 ที่ได้รับการดัดแปลงพันธุกรรมซึ่งมีพลาสมิดรีคอมบิแนนท์ pGEX (CGRP/msCT) ที่ให้ผลบวก ถูกนำไปเพาะเลี้ยงในอาหารเหลว LB ที่มีแอมพิซิลลิน 50 μg·mL⁻¹ และเพาะเลี้ยงข้ามคืนที่อุณหภูมิ 37°C จากนั้น นำเชื้อที่ได้มาเพาะเลี้ยงเพิ่มจำนวนในขวดอาหารเหลว LB สดใหม่ขนาด 50 มล. จำนวน 10 ขวด ที่มีแอมพิซิลลิน 50 μg·mL⁻¹ ในอัตราส่วน 1:100 และเมื่อค่า OD600 อยู่ในช่วง 0.6-0.8 จึงเติม IPTG จนได้ความเข้มข้นสุดท้าย คือ 0.1, 0.2, 0.3, 0.4, 0.5, 0.6, 0.7, 0.8, 0.9, 1.0 mmol·L⁻¹ หลังจากกระตุ้นการเจริญเติบโตของแบคทีเรียที่อุณหภูมิและเวลาเดียวกันแล้ว นำสารละลายแบคทีเรียปริมาณ 1 มิลลิลิตร มาแยกเซลล์แบคทีเรียโดยการปั่นเหวี่ยง และนำไปวิเคราะห์ด้วย SDS-PAGE เพื่อศึกษาอิทธิพลของความเข้มข้นของ IPTG ที่แตกต่างกันต่อการแสดงออกของโปรตีน จากนั้นเลือกความเข้มข้นของ IPTG ที่ให้การแสดงออกของโปรตีนมากที่สุด
จากการทดลองพบว่า ความเข้มข้นของ IPTG ไม่ควรสูงเกินไป เนื่องจาก IPTG มีความเป็นพิษต่อแบคทีเรีย การใช้ความเข้มข้นสูงเกินไปจะทำให้เซลล์ตายได้ และโดยทั่วไปแล้ว เราหวังว่ายิ่งมีโปรตีนที่ละลายน้ำได้ในเซลล์มากเท่าไหร่ก็ยิ่งดี แต่ในหลายกรณี เมื่อความเข้มข้นของ IPTG สูงเกินไป จะทำให้เกิดสารตกค้างจำนวนมาก แต่ปริมาณโปรตีนที่ละลายน้ำได้กลับลดลง ดังนั้น ความเข้มข้นของ IPTG ที่เหมาะสมที่สุดจึงไม่ใช่ยิ่งสูงยิ่งดี แต่เป็นยิ่งต่ำ
จุดประสงค์ของการชักนำและการเพาะเลี้ยงสายพันธุ์ที่ได้รับการดัดแปลงทางพันธุกรรมคือการเพิ่มผลผลิตของโปรตีนเป้าหมายและลดต้นทุน การแสดงออกของยีนเป้าหมายไม่เพียงแต่ได้รับผลกระทบจากปัจจัยของสายพันธุ์เองและพลาสมิดที่ใช้ในการแสดงออกเท่านั้น แต่ยังได้รับผลกระทบจากสภาวะภายนอกอื่นๆ เช่น ความเข้มข้นของสารชักนำ อุณหภูมิในการชักนำ และระยะเวลาในการชักนำ ดังนั้น โดยทั่วไปแล้ว ก่อนที่จะทำการแสดงออกและทำให้บริสุทธิ์ของโปรตีนที่ไม่รู้จัก ควรศึกษาเวลา อุณหภูมิ และความเข้มข้นของ IPTG ในการชักนำ เพื่อเลือกสภาวะที่เหมาะสมและได้ผลการทดลองที่ดีที่สุด
วันที่โพสต์: 31 ธันวาคม 2021